

本文原文为:Resistant starch intake facilitates weight loss in humans by reshaping the gut microbiota
本文原作者Huating Li,教授,博士生导师,国家自然科学基金优秀青年基金获得者。现就职于上海交通大学附属第六人民医院内分泌科。主要研究方向为糖尿病发病机制及诊治新靶点。
新的证据表明,膳食纤维调节肠道微生物群可能为代谢紊乱提供解决方案。在一项针对 37 名超重或肥胖参与者的随机安慰剂对照交叉设计试验 (ChiCTR-TTRCC-13003333) 中,作者测试了抗性淀粉 (RS) 作为膳食补充剂是否会影响肥胖相关的结果。
结果显示,补充 RS 8 周有助于实现体重减轻(平均 -2.8 公斤)并改善体重超重个体的胰岛素抵抗。RS 的好处与肠道微生物群组成的变化有关。补充青春双歧杆菌 (一种与减轻研究参与者肥胖显着相关的物种)可以保护雄性小鼠免受饮食引起的肥胖。
从机制上讲,RS 诱导的肠道微生物群变化会改变胆汁酸分布,通过恢复肠道屏障来减少炎症并抑制脂质吸收。我们证明,RS 至少可以部分通过青春双歧杆菌促进体重减轻,并且肠道微生物群对于 RS 的作用至关重要。
01
综述
当前全球肥胖流行,研究的方向是新的肥胖预防和减肥策略。这一点至关重要,因为肥胖会显着导致糖尿病和心血管疾病等合并症,而这些疾病是全球主要的死亡原因。相反,减肥可以减轻这些合并症,强调体重管理在预防和治疗这些疾病方面的重要性。
肠道微生物群越来越被认为是宿主生理学和病理生理学的重要调节因子。具体来说,之前的研究报道肠道微生物群调节炎症、脂肪储存和葡萄糖代谢。尽管从健康捐献者到肥胖个体的粪便微生物移植 (FMT) 的结果不一致或短期,但将饮食干预与 FMT 相结合可能会导致受者微生物群发生有利的改变并改善临床结果。因此,通过饮食干预合理控制肠道微生物组可能是一种有前途的抗肥胖策略。
益生元,包括多糖、寡糖和其他可发酵膳食纤维,可增加有益肠道微生物群的数量,特别是某些双歧杆菌和乳酸菌 。这些细菌可以减少病原体数量、强化肠道屏障并减轻炎症反应。
此外,可以定制用不同纤维制剂配制的零食来改变与微生物组特定元素相关的功能;然而,大多数益生元研究都是基于相关性,而没有建立肠道微生物群调节与观察到的对新陈代谢的有益影响之间的因果关系。来自对限生动物的人体试验和机制研究的见解对于确定微生物组改变与宿主生物反应之间的因果关系至关重要。这些研究对于理解微生物组变化与益生元或其他可发酵膳食纤维的生理优势之间的联系机制至关重要。
RS是指一种可发酵膳食纤维,它不能被小肠中的人类淀粉酶消化,并进入结肠,在结肠中被肠道微生物群发酵。对啮齿动物的研究表明,与可消化淀粉喂养相比,RS 可以导致全身脂肪减少,特别是内脏脂肪。当碳水化合物成分由小鼠的 RS 组成时,低蛋白质和高碳水化合物的饮食会产生最有利的代谢结果;然而,人类数据表明,在服用 RS 4 至 12 周后,代谢综合征患者的总体重没有受到影响。
补充 RS 的低脂饮食对宿主具有有益作用,但高脂肪饮食会减弱 RS 发酵和有益作用。这可能是为什么在上述临床试验中 RS 似乎对体重没有影响的一种可能解释,因为这些临床试验对饮食的依从率不高。此外,这意味着 RS 相关的肠道微生物群在 RS 的治疗效果中发挥着至关重要的作用。然而,RS 作为一种功能性、适应性强的食品成分用于人类肥胖治疗的潜力以及 RS 相关肠道微生物组改变对代谢益处的调节仍不清楚。
因此,在肥胖个体中进行强有力的试验对于证实 RS 对消费者不同生理方面的影响以及所需剂量的主张至关重要。此外,应使用多组学方法和限生动物模型来系统地、机械地连接RS对肠道微生物群落和宿主代谢的影响。
作者对体重超重的个体进行了一项交叉随机临床试验,以研究 RS 作为膳食补充剂对肥胖和其他代谢表型的影响。该试验是一项提供等能量和平衡背景饮食的喂养研究。进行宏基因组学和代谢组学分析以评估 RS 对肠道微生物群组成和功能的影响。
此外,作者分析了从选定的人类供体转移到抗生素治疗小鼠的 RS 修饰的肠道微生物群对宿主肥胖和葡萄糖代谢的影响,还探讨了肠道微生物群通过 RS 赋予代谢优势的机制。
02
结论
RS干预有利于减肥
该调查是一项安慰剂对照、双盲、交叉设计干预措施 (ChiCTR-TTRCC-13003333),涉及 37 名参与者(平均年龄 33.43 ± 7.71 岁)。参与者的体重指数(BMI)≥24 kg m −2和/或腰围增加(男性≥85 cm,女性≥80 cm)。没有人患有慢性疾病、正在接受影响葡萄糖代谢的治疗或最近使用过抗生素或益生菌(三周内)。
为期 20 周的研究持续时间包括两个为期 8 周的干预期,其中高直链淀粉玉米 (HAM-RS2)(RS,2.8 kcal g −1 , 91.2 g,含 40 g RS)和对照淀粉 (CS) 各干预一个干预期(AMIOCA)(3.55 kcal g -1,72 g,支链淀粉,含有0 g RS,能量供应相等)和干预之间4周的冲洗期(图1a)。
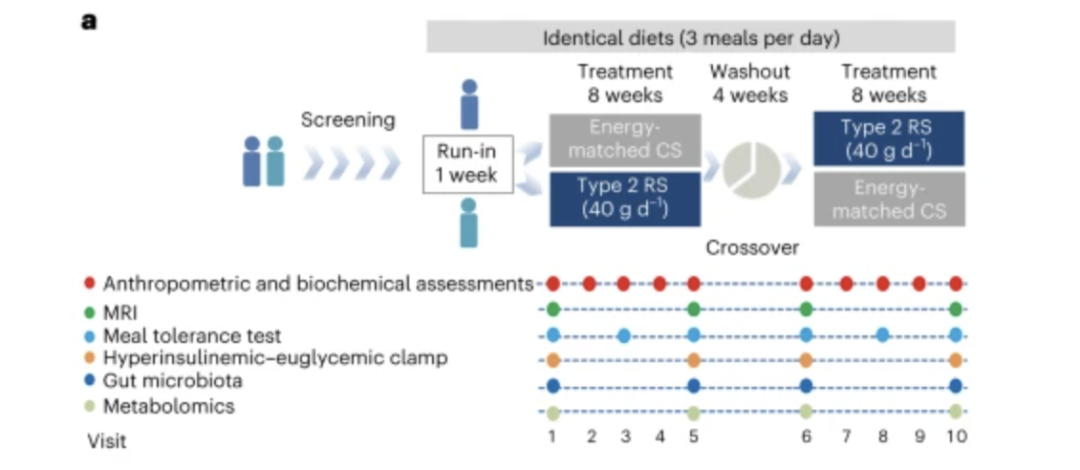
图1a:临床试验图。在入组、随机化和磨合期之后,参与者交替服用 RS 或 CS,并间隔一个洗脱期。在整个试验期间,为所有参与者提供相同的饮食。每次访问的评估都显示在图表中
研究参与者被随机分为两组:(1) RS-Washout-CS 或 (2) CS-Washout-RS。淀粉以粉末形式装在预包装袋中,与 300 毫升水混合。每位参与者每天两次,饭前 10-15 分钟服用一袋。在整个试验过程中,包括磨合期、两次干预和干预冲洗期,我们根据中国和美国成人超重和肥胖预防和管理指南提供等能量和均衡的背景饮食(一日三餐)20 , 21.RS 和 CS 干预之间,参与者在不进餐或淀粉消耗方面没有差异,这表明 RS 和 CS 干预之间的饮食依从性没有差异。除了膳食纤维摄入量的差异(RS与CS,53.84±4.70克与9.97±5.17克,P <0.001)外,RS或CS干预期间消耗的总能量和大量营养素的百分比相似(补充表1)。总共有 37 名参与者,包括 22 名男性和 15 名女性参与者完成了研究并被纳入分析(表1和扩展数据图1)。该研究于2013年7月3日至2016年10月14日在中国上海进行,没有报告胃肠道副作用,如恶心、呕吐、腹胀、排便增加或大便频率改变。
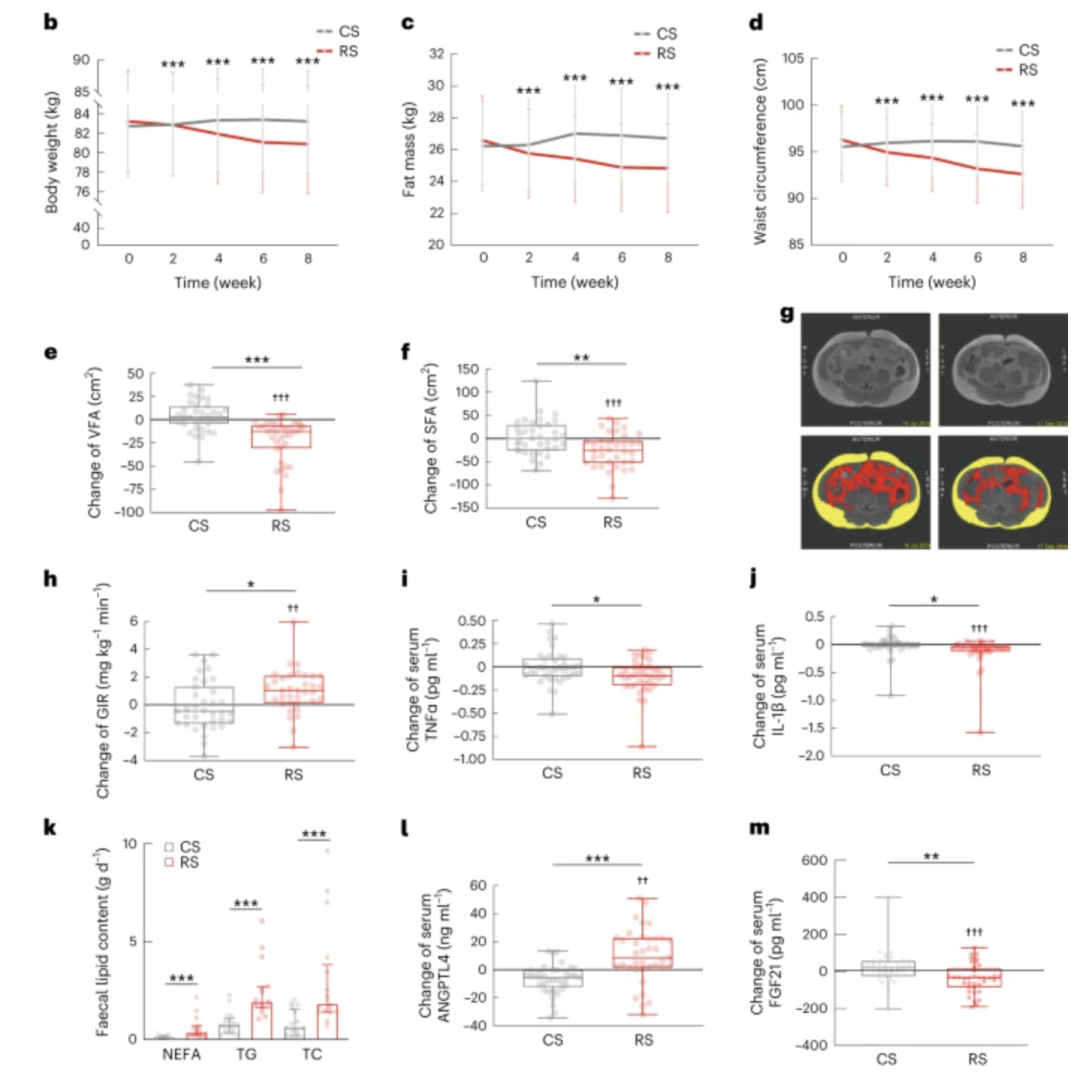
图1b-m:b-d,RS干预显着降低了体重(b)、脂肪量(c)和腰围(d)。e、f,通过 MRI 评估 VFA 和 SFA 的变化。g,参与者在 8 周 RS 干预之前(左)和之后(右)的代表性腹部 MRI。肚脐水平的原始(上)和标记(下)MRI。黄色代表SFA,红色代表VFA。h,通过高胰岛素-正常血糖钳夹评估的 GIR 变化。i,血清TNFα水平的变化。j,血清IL-1β水平的变化。k,RS或CS干预8周后每日粪便脂质排泄,包括NEFA、TG和TC。l、血清ANGPTL4水平的变化。m,血清FGF21水平的变化。对于 RS 或 CS,n = 37 个人 ( b – d , j , l , m ),n = 36 个人 ( e , f , i ),n = 35 个人 ( h ) 和n = 17 个人 ( k )。通过基线值调整的协方差分析 (ANCOVA) 用于在每次访问 ( b – d )时比较 RS 和 CS 。数据显示为平均值(95% 置信区间 (CI))。*** P < 0.001。数据显示为带有 IQR ( k ) 的中位数。非参数 Wilcoxon 秩和检验用于评估两种干预措施之间的显着性。*** P < 0.001。数据显示为盒须图(e、f、h – j、l、m)。箱线图、中位数和四分位数;晶须,数据范围。* P = 0.025、0.014 和 0.046 ( h – j ),** P = 0.004 和 0.002 ( f , m ),*** P < 0.001 为根据干预顺序调整的线性混合模型评估的组间差异。†† P = 0.003 和 0.002 (小时,l)。††† 通过混合线性模型调整干预后的 Bonferroni 检验,组内变化P < 0.001。
我们使用按干预顺序调整的线性混合模型比较了干预之间的人体测量参数和生化指标的实际变化(RS与CS)以及干预前后的组内变化(表1和补充表2 )。RS干预后主要结局体重显着下降,RS干预后相对于CS干预的净绝对变化为-2.81 kg(95% CI -3.55 kg至-2.07 kg;P < 0.001),而RS干预后体重没有显着变化。CS干预后观察到(图1b)。
此外,与CS干预相比,RS干预后脂肪量和腰围显着减少(图1c,d)。在RS干预期间,参与者的体重、腰围和脂肪量从第2周开始显着下降。通过腹部磁共振成像 (MRI) 测量的内脏脂肪面积 (VFA) 和皮下脂肪面积 (SFA) 与 CS 消耗后相比,RS 消耗后的内脏脂肪面积 (VFA) 和皮下脂肪面积 (SFA) 均较低(分别为P < 0.001 和P = 0.004;图1e -G)。无论是RS-Washout-CS组还是CS-Washout-RS组,RS干预后体重和其他肥胖相关结果均显着下降(均P < 0.05)。在 RS-Washout-CS 组中,肥胖相关结果显示在清除期后恢复至基线水平(补充表3和扩展数据图2a-e)。此外,双向方差分析(ANOVA)显示干预后体重和肥胖相关结果(脂肪量、腰围和 VFA)存在显着差异(P < 0.001),并且没有显着的顺序效应或干预顺序交互作用。这些结果表明,为期 8 周的 RS 干预可减少体重超标个体的腹部肥胖。
此外,RS干预后葡萄糖耐量显着改善(扩展数据图2f,g)。RS干预后参与者在餐耐量测试(MTT)后120分钟时的胰岛素浓度显着低于CS干预后(扩展数据图2h);然而,与基线值相比,CS 干预并没有引起葡萄糖和胰岛素水平的任何差异(扩展数据图2f,h)。我们进一步通过高胰岛素-正常血糖钳夹评估胰岛素敏感性,发现与CS相比,RS干预后葡萄糖输注率(GIR)显着增加(中位增加1.05 mg kg -1 min -1 , 0.15至2.10)干预(P = 0.025)(图1h),表明胰岛素敏感性显着改善。此外,RS干预后观察到血清脂联素水平显着增加(扩展数据图2i)。此外,RS干预后,第一相和第二相胰岛素分泌与CS干预后均无显着差异(补充表4)。所有这些结果表明,为期 8 周的 RS 干预改善了体重超重个体的葡萄糖耐量和胰岛素敏感性。
为了研究 RS 促进减肥的潜在机制,我们确定了与肥胖密切相关的慢性、低度炎症反应和肠道脂质消化的变化。研究发现,与食用 CS 相比,食用 RS 后的研究参与者中血清肿瘤坏死因子 (TNF)α 和白细胞介素 (IL)-1β 等促炎细胞因子的水平显着降低(分别为 P = 0.014 和P = 0.046)。尽管在RS和CS消耗期间单核细胞趋化蛋白-1(MCP-1)、IL-10和IL-6没有观察到显着差异(图1i、j和表1)。
此外,我们测量了研究参与者的粪便脂质,发现与食用CS相比,食用RS后粪便中非酯化脂肪酸(NEFA)、甘油三酯(TG)和总胆固醇(TC)的每日排泄量显着更高(图1)。
由于RS和CS消耗期间脂肪摄入量没有显着差异(补充表1),这些结果表明RS干预可能会减少饮食中的脂质吸收。与摄入CS相比,摄入RS后,研究参与者体内血管生成素样4(ANGPTL4)的循环水平显着升高,血管生成素样4(ANGPTL4)是肠道与脂质代谢之间的潜在联系3 (图1l)。据报道,血清成纤维细胞生长因子 21 (FGF21) 在肥胖状态下会增加,但在食用 RS 后显着降低(图1m)。
RS 干预重塑肠道微生物群
为了研究干预期间肠道微生物群的动态,进行了鸟枪法宏基因组测序。每个样本的平均吞吐量为 5.74 (sd 0.87) Gbp。基于样本平均丰度前 50 个物种的物种级分类学特征显示,普氏 菌、粪类 杆菌和普氏粪 杆菌是样本中最常见的。大多数样本中其他物种的丰度要低得多(每个物种平均为 1.1%)(扩展数据图3a)。
为了评估肠道微生物组的组成因 RS 干预而改变的程度,我们根据样本之间的物种变异概况(8 周干预后丰度变化倍数)计算了 Bray-Curtis 距离使用以下步骤。物种变异概况由每个物种处理后丰度的log 2倍变化 (log 2 FC) 组成。倍数变化值标准化为 0 到 1,每个样本中的总和为 1。我们根据物种变异的概况进一步计算了布雷-柯蒂斯距离。基于Bray-Curtis距离的非度量多维标度(NMDS)图显示,RS和CS样本在一定程度上分离,表明RS和CS干预后微生物群的变化模式分别存在显着差异(P < 0.001(来自 VEGAN 的 Adonis 测试)(图2a)。
因此,这一发现表明 RS 干预会影响肠道微生物群的动态并重组其组成。CS 和 RS 干预基线之间的微生物组成没有显着差异(扩展数据图3b-e)。我们发现七个物种 与整体变异概况(对于所有 RS 和 CS 样本)显着相关(Benjamini-Hochberg (BH) 错误发现率 (FDR) 调整后P < 0.01)(图2a)。我们发现BMI与总体分类变异谱显着相关(P < 0.001,R包VEGAN中的envfit测试)(图2a),表明体重的变化与重组的肠道微生物组相关。
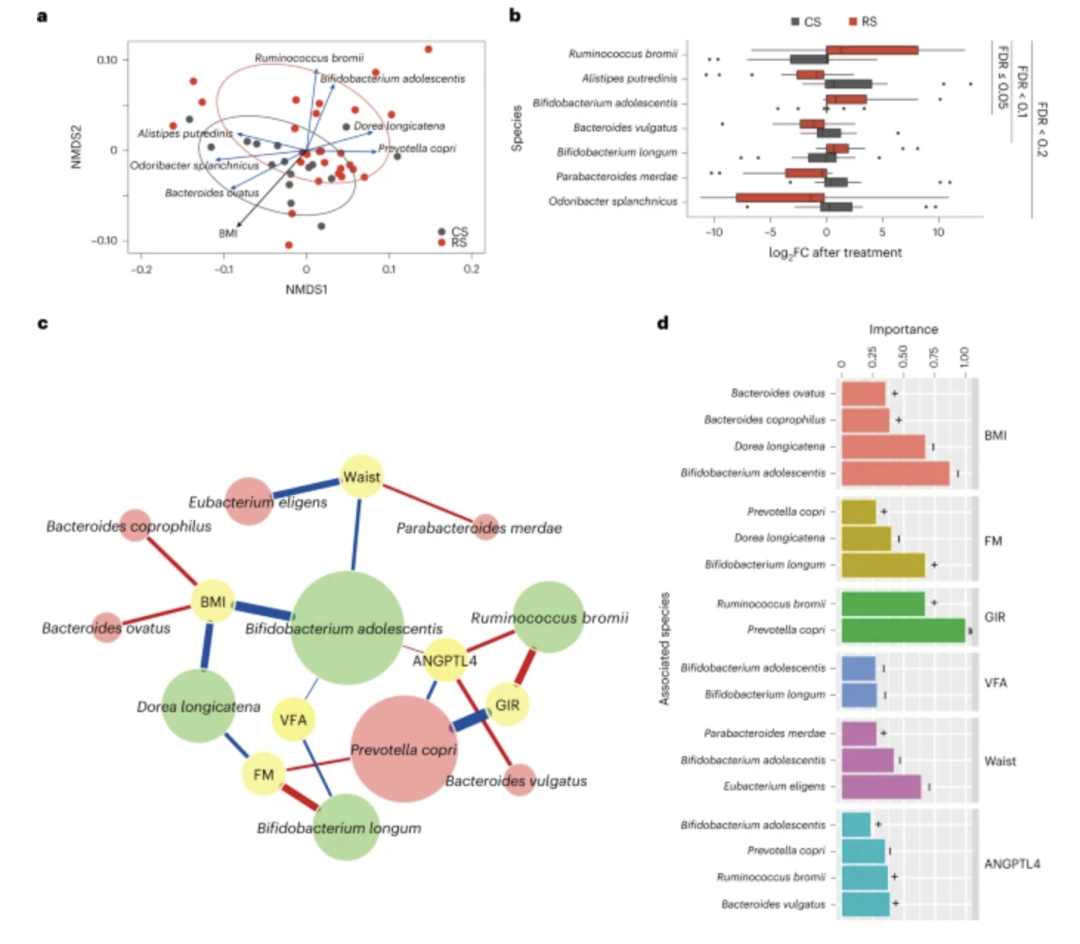
图 2:RS 治疗后肠道微生物群与宿主表型的关联
a,基于物种丰度变化(干预后倍数变化)的样本 NMDS。红色和灰色圆圈分别围绕(95% CI)RS 和 CS 样本。与总体排序显着相关的物种和表型(FDR 使用 VEGAN R 包中的 envfit 函数调整P < 0.01)用箭头突出显示(蓝色,物种;黑色,表型),其中箭头的长度反映了关联的强度。b,RS 和 CS 干预之间有 7 个物种具有显着不同的变异特征(P < 0.05,BH FDR < 0.2,配对两侧 Wilcoxon 符号秩检验)。箱线图显示中位数和 IQR。晶须延伸至 1.5 × IQR。c,肠道微生物与宿主表型之间的关联网络。RS 组和 CS 组之间共有 12 个具有不同变异(P < 0.05,BH FDR < 0.3,双边 Wilcoxon 秩和检验)的物种被用作回归模型中的预测因子。节点颜色反映了响应 RS 的表型(黄色)、丰度增加的物种(绿色)或丰度减少的物种(红色)。连接线的颜色反映了微生物与表型之间的正相关(红色)或负相关(蓝色)。连接线的宽度反映了丰度变异(干预后倍数变化)和表型变异(干预后倍数变化)之间统计线性相关性(模型平均重要性)的强度。重要性是模型选择过程中包含此类变量的广义线性模型的 AIC 权重之和。物种的节点大小反映了其对所有表型的总体影响。d,每个表型相关物种重要性的总结。在a – d中,从所有参与者中随机选择 RS 前后 27 名个体和 CS 前后 16 名个体进行宏基因组分析。
为了确定与 RS 干预相关的肠道微生物群特征,我们比较了 RS 和 CS 处理后每个物种丰度的变化(作为 8 周干预后丰度的倍数变化)。
结果显示,青春双歧杆菌 、长双歧杆菌和布氏瘤胃球菌3个菌种在RS干预后显着增加,而在CS干预后保持稳定或下降。相反,RS 处理后,包括腐臭拟杆菌、普通拟杆菌、内脏恶臭杆菌和Merdae副杆菌在内的四个物种在 RS 处理后减少(P < 0.05,BH FDR < 0.2,Wilcoxon 符号秩检验),但在 CS 处理后保持稳定或增加。其中,与CS干预相比, RS干预后R. bromii、A. putredinis和B.teenis是变化最显着的物种(FDR = 0.05)(图2b和补充表5)。
为了确定与 RS 干预益处相关的物种,我们研究了 12 个物种(通过 Wilcoxon 秩和检验 FDR < 0.3)与宿主的六个显着改变的参数的相关性,包括 BMI、脂肪量、腰围、VFA、GIR 和血清ANGPTL4 水平。广义线性模型量化了物种丰度变化与表型变化之间的关联。物种和表型之间关联的强度是通过线性模型中变量的重要性来描述的,该线性模型基于使用校正的赤池信息标准(AIC)进行的模型选择,该标准量化了用于给定的统计模型的相对质量/权重数据。
一个变量的重要性反映了所有可能的加权线性模型的总体支持度或包含该变量的所有可能的线性模型总权重的比例。在具有差异丰度的12个物种中,青春 双歧杆菌与各种表型(与表型的链接总数)具有最频繁的关联(图2c)。
在表型中,BMI和GIR与肠道微生物的关联性最强(相关物种的平均重要性> 0.5)(图2d)。在单一物种水平上,我们发现补充 RS 后R. bromii丰度的增加与 GIR 的增加相关(重要性为 0.67,P < 0.001)。结合上述R. bromii的显着增加,我们的研究结果表明R. bromii可能是响应 RS 干预的关键物种,这与之前的研究一致。
青春 双歧杆菌可能在缓解肥胖方面发挥着至关重要的作用,其与较低的 BMI、腰围和 VFA 的强相关性就证明了这一点(重要性分别为 0.87、0.47 和 0.27;P < 0.001)。其丰度增加还与较高的血清ANGPTL4水平呈正相关(重要性0.252,P <0.001),表明潜在的脂质代谢作用。作为主要的 RS 降解者,我们探讨了R. bromii和B.publishis 在基线时的存在及其与关键结果的关系。RS干预后,青春 双歧杆菌阳性个体的脂肪量下降更多,ANGPTL4水平增加更多(均P < 0.05),表明肠道菌群的初始组成,尤其是青春 双歧杆菌,与获益密切相关。在RS和CS干预之间丰度差异最大的十大代谢相关京都基因和基因组百科全书(KEGG)途径中,亚油酸代谢(ko00591)和双酚降解(ko00363)途径在RS处理期间减少,而它们在 CS 处理中显着增加(两种途径的 FDR = 0.057)。
RS改变的肠道微生物群与代谢物的关联
虽然尚不清楚肠道微生物群的变化如何为宿主带来益处,但可能的机制是通过改变代谢产生。因此,我们进一步对研究参与者的血清进行了非靶向代谢组学分析。
与CS干预相比,RS干预后与肥胖相关的血清代谢物,如肉碱和蛋氨酸降低(补充表7)。我们使用广义线性模型来研究 RS 干预期间肠道微生物与血清代谢物变化之间的关系。RS干预期间青少年双歧 杆菌的变化与血清肉碱、能量代谢和脂质代谢物的变化呈负相关(平均重要性0.15;Fisher组合P <0.05;扩展数据图6a)。相反,RS干预期间普通双歧杆菌的变化与能量代谢代谢物的变化呈正相关,包括黄嘌呤和尿酸。
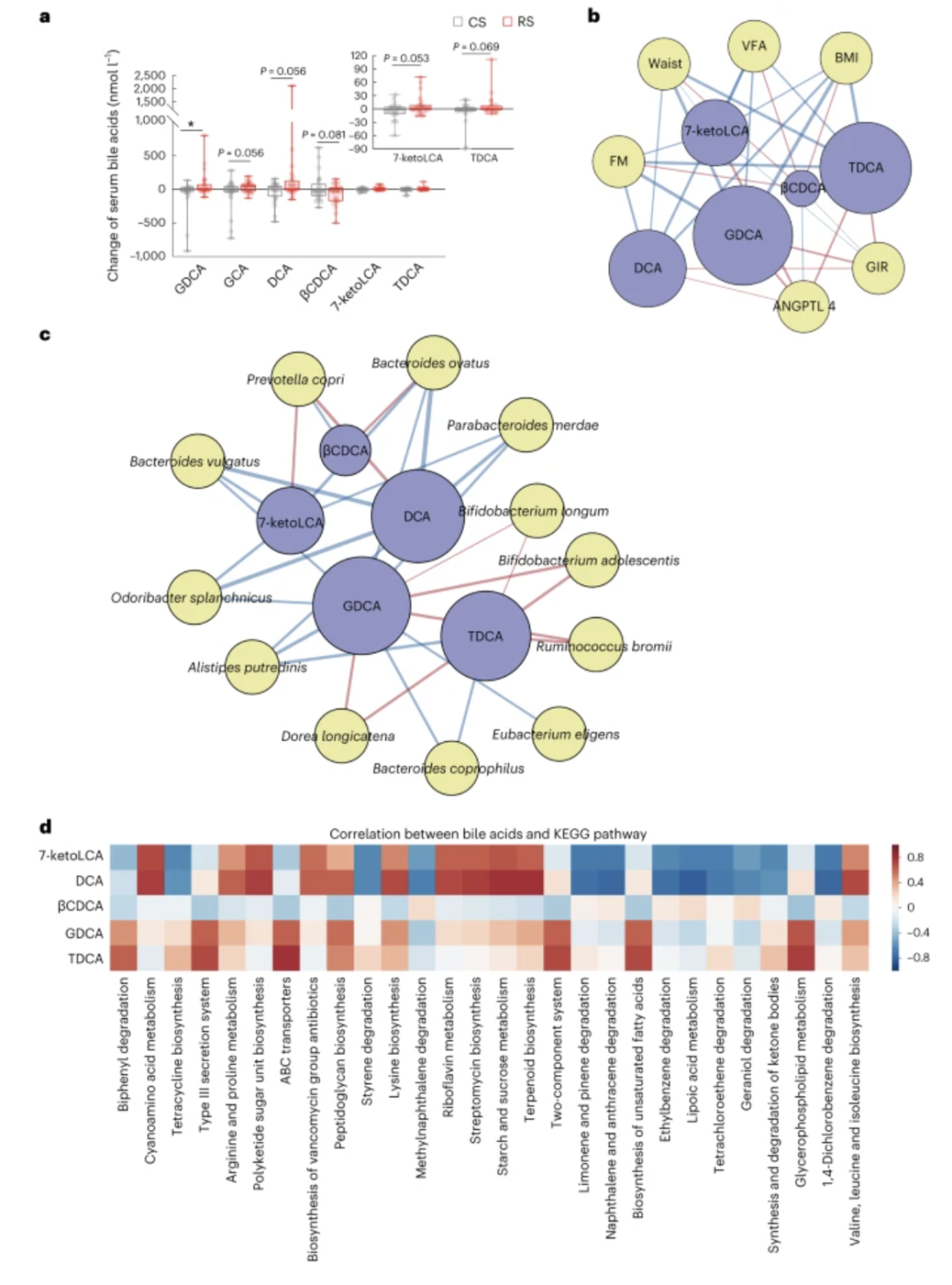
图 3:胆汁酸介导肠道微生物组和宿主表型之间的相互作用
a,8 周 RS 或 CS 干预后评估参与者血清胆汁酸水平的倍数变化。数据以盒须图的形式呈现。每个框中间的水平线表示中值,框的顶部和底部边框表示第 75 个和第 25 个百分位数,胡须表示最低值和最高值。* 混合线性模型P = 0.036。b,胆汁酸与宿主表型之间的关联网络。c,肠道微生物与宿主表型之间的关联网络。节点颜色反映胆汁酸(蓝色)、表型(b中的黄色)或微生物(c中的黄色)。连接线的颜色反映了胆汁酸与表型之间或胆汁酸与微生物之间的正相关(红色)或负相关(蓝色)。连接线的宽度反映了胆汁酸变化(干预后倍数变化)与表型变化(干预后倍数变化)之间或丰度变化(倍数变化)之间统计线性关联(模型平均重要性)的强度。干预后)和胆汁酸的变化(干预后倍数变化)。重要性是模型选择过程中包含此类变量的广义线性模型的 AIC 权重之和。胆汁酸的节点大小反映了其对所有表型或微生物的总体影响。d,KEGG通路分析显示胆汁酸的变化与KEGG通路丰度变化之间的相关性。
由于胆汁酸是将肠道微生物群与宿主联系起来的重要信号代谢物,我们对胆汁酸谱进行了定量分析。甘氧胆酸 (GDCA) 是唯一一种在 RS 和 CS 干预之间具有统计学显着差异的胆汁酸。其他胆汁酸,如甘胆酸(GCA)、脱氧胆酸(DCA)、7-酮石胆酸(7-ketoLCA)和牛磺脱氧胆酸(TDCA)表现出增加的趋势,而3β-鹅脱氧胆酸(βCDCA)水平则呈下降趋势。RS干预后减少。
通过相关性分析发现,次级胆汁酸GDCA和TDCA与代谢表型的相关性最密切。GDCA和TDCA的变化与肥胖相关表型呈负相关,而与胰岛素敏感性和ANGPTL4水平呈正相关(图3b)。RS处理后GDCA和TDCA的变化与青春双歧杆菌、长双歧杆菌、Dorea longicatena和R. bromii等4个物种呈显着正相关,而 与另外8 个物种呈 负相关, 包括腐烂放 线菌、嗜粪拟 杆菌、卵形拟杆菌( Bacteroides ovatus)、普通拟杆菌( B. vulgatus)、真杆菌(Eubiotics eligens)、内脏O. splanchnicus、P. merdae和P. copri(图3c)。此外,我们还发现了一些与几种代谢物的变化密切相关的微生物途径,包括GDCA、TDCA、βCDCA、DCA和7-ketoLCA(图3d)。我们的结果表明,RS 影响的肠道微生物群增加了次级胆汁酸的产生,这可能会减轻肥胖和胰岛素抵抗。为了确定该物种是否与胆汁酸代谢相关,我们研究了肠道微生物组与胆汁盐水解酶(BSH;K01442)之间的关联,胆汁盐水解酶是胆汁酸代谢和宿主-微生物组串扰的看门人31。我们的结果显示,与 CS 干预相比,RS 干预中肠道微生物群落水平内BSH基因的丰度显着降低( P < 0.05,Wilcoxon 秩和检验)。我们发现BSH的表达与B. vulgatus显着相关(r = 0.561,P < 0.001;补充表9)。
短链脂肪酸 (SCFA) 主要由膳食纤维的肠道微生物发酵产生,在维持肠道和代谢健康方面发挥着重要作用28。因此,我们进行了靶向代谢组学来研究 RS 和 CS 治疗后粪便和循环 SCFA 的变化。我们发现,与 CS 干预相比,RS 干预后,粪便中异丁酸和戊酸的浓度显着降低,而其他粪便 SCFA,包括乙酸盐、丙酸盐、丁酸盐和己酸盐,在 RS 干预后没有显着差异(扩展数据图6b-g))。RS 和 CS 干预之间的 SCFA 循环水平没有显着差异。
RS 重塑肠道微生物群减轻小鼠肥胖
为了研究 RS 相关肠道微生物群改善宿主腹部肥胖和葡萄糖代谢的潜力,我们在接受抗生素治疗的西式饮食小鼠中进行了 FMT,利用 RS 或 CS 干预后来自人类捐赠者的样本(其中干预后体重接近各组平均值; 每组n = 4)(图4a)。我们观察到供体和接受 FMT 的小鼠之间 RS 诱导的变化趋势一致(扩展数据图7a)。FMT两周后,接受RS微生物群的小鼠的体重和脂肪量低于接受CS微生物群的小鼠(P <0.001)(图4b,c)。RS菌群定植的小鼠附睾白色脂肪组织(eWAT)、肾周白色脂肪组织(pWAT)和肠系膜白色脂肪组织(mWAT)的储存质量百分比显着低于CS菌群定植的小鼠(图.4d ,e )。eWAT 和 mWAT 的组织学分析显示,在 RS 微生物群定植的小鼠中,脂肪细胞的大小显着减小(图4f,g)。
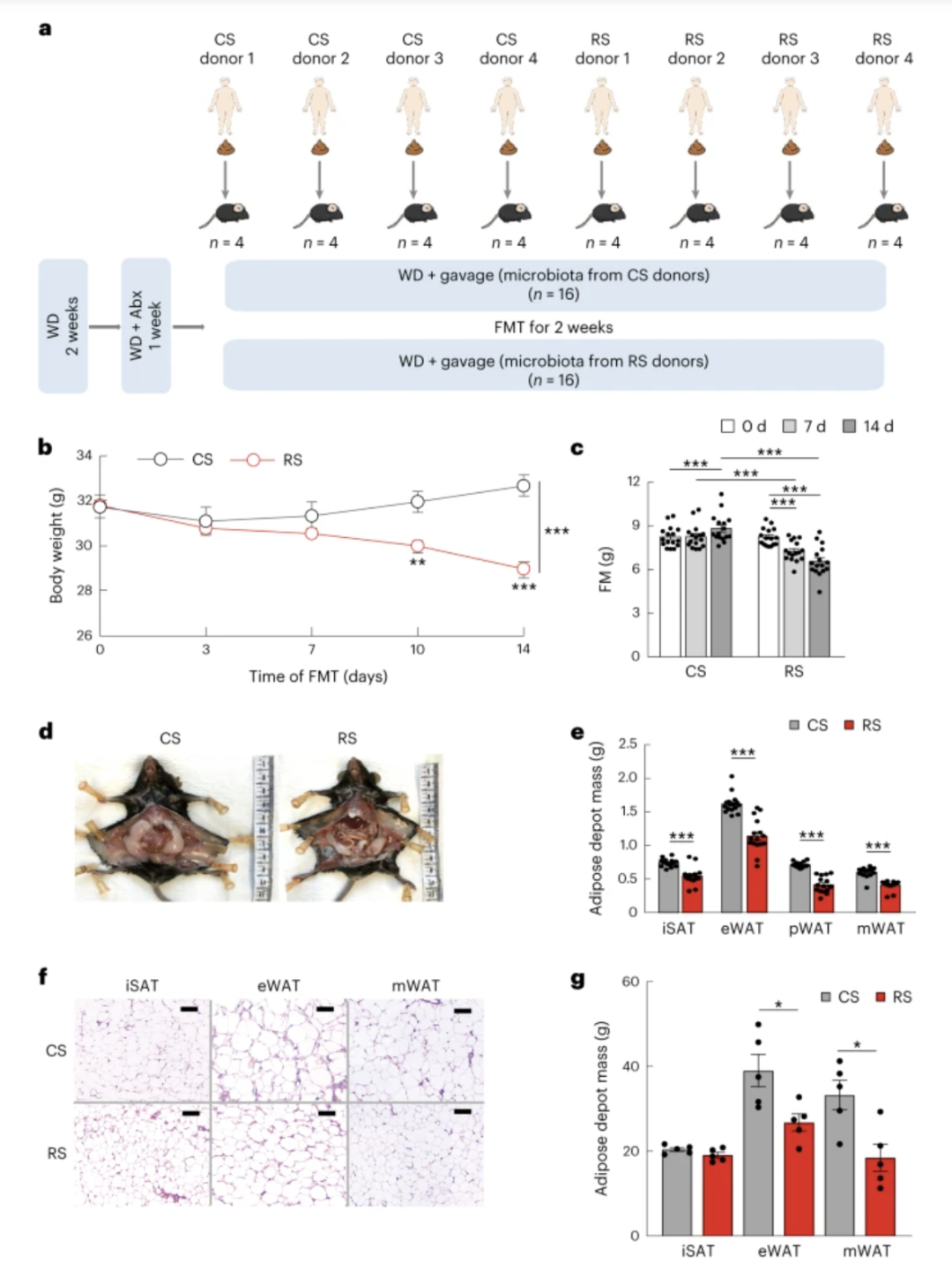
图 4:RS 影响的肠道微生物群可减轻肥胖
a,FMT示意图。经过 8 周 RS 或 CS 干预(每次干预n = 4)后,人类捐赠者的粪便样本被移植到抗生素 (Abx) 治疗的 C57BL/6 小鼠体内。在定植前 2 周以及定植期间的 14 天期间,给小鼠喂食经过辐照的西方饮食 (WD)( 每组n = 16)。b、c、FMT 后体重和脂肪量的变化。d,来自 RS 或 CS 供体的微生物群定植 14 天的小鼠内脏脂肪的代表性照片。e,微生物群定植 14 天的小鼠体内的脂肪库质量,包括腹股沟皮下脂肪 (iSAT)、eWAT、pWAT 和 mWAT。f、g,两组定植有 CS(上)或 RS 微生物群(下)的小鼠 iSAT、eWAT 和 mWAT 的苏木精和伊红染色切片的代表性图像和定量数据。比例尺,100 µm。数据在三个独立实验中重现。数据以平均值±sem表示。显着性由配对双尾学生t检验(c)和不配对双尾学生t检验(b,e,g)(正态分布)或非参数两侧Wilcoxon等级确定。总和检验(非正态分布)。* P = 0.02 和 0.01 ( g ), ** P = 0.002 ( b ), *** P < 0.001 ( b , c , e )。
综合实验动物监测系统表明,两组小鼠在能量消耗、呼吸交换比、CO 2产生、O 2消耗、活动和食物摄入方面没有表现出显着差异(扩展数据图7b-g)。在 RS 干预后,通过移植人类肠道微生物群,这些小鼠的葡萄糖耐量至少得到部分改善(扩展数据图7h,i)。RS微生物群定植的小鼠中的循环脂联素水平显着高于CS微生物群定植的小鼠(扩展数据图7j)。这些结果表明,RS 诱导的肠道微生物群变化足以缓解肥胖并改善宿主葡萄糖代谢。
RS 重塑肠道微生物群恢复肠道屏障
与临床试验结果一致,RS 改变的肠道微生物群改善了全身炎症。FMT两周后,RS菌群定植的小鼠中循环MCP-1、IL-1β和IL-6水平显着低于CS菌群定植的小鼠(均P <0.01)(图5a)。RS微生物群定植的小鼠中抗炎因子IL-10的血清水平增加(图5a)。在肠系膜脂肪组织中,与接受CS微生物群的小鼠相比,接受RS微生物群的小鼠中MCP-1、IL-1β和IL-6的表达也显着较低,而IL-10的表达较高(图5b)。
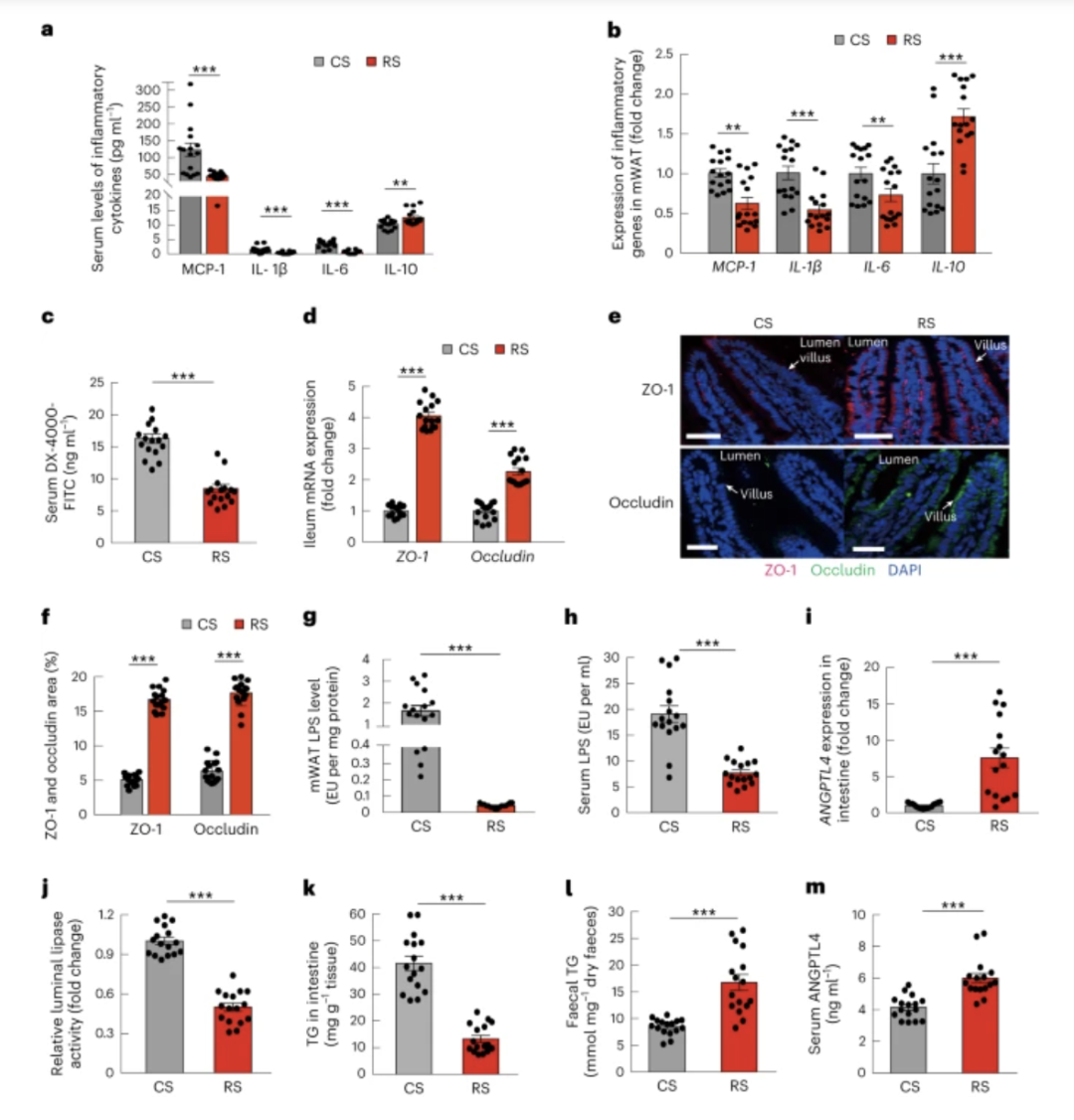
图 5:RS 影响的肠道微生物群恢复肠道屏障并减少脂质吸收
小鼠如图4所示进行分组和治疗( 每组n = 16)。a,来自 RS 或 CS 供体的微生物群定植的小鼠中炎症细胞因子的血清水平。b,mWAT 和来自 RS 或 CS 供体的微生物群定植的小鼠中炎症基因的表达。c,体内肠道通透性。d ,回肠中ZO-1和occludin的表达。e,通过免疫荧光和用 4,6-二脒基-2-苯基吲哚 (DAPI)(蓝色)复染观察肠绒毛中 ZO-1(红色)和 occludin(绿色)的定位和水平。显示了每组的代表性图像(比例尺,50 µm)。f,通过ImageJ软件对ZO-1和occludin的阳性染色面积进行定量分析,并计算为总病灶面积的百分比。g、h、mWAT 和循环(血清)中的 LPS 水平。i,回肠中ANGPTL4的表达水平。j,相对肠腔脂肪酶活性。k,回肠中的TG水平。l,粪便TG水平。m,血清ANGPTL4水平。数据在三个独立实验中重现。数据以平均值±sem表示。显着性通过不配对的双尾学生t检验(正态分布)或非参数两侧Wilcoxon秩和检验(非正态分布)来确定。** P = 0.005 ( a ) 和 0.001 和 0.010 ( b ),*** P < 0.001。
代谢性内毒素血症可能是由于脂多糖 (LPS) 从肠道到循环系统的渗透增加所致,是肥胖引起的慢性炎症的关键因素。
为了研究 RS 改变的肠道微生物群如何与代谢性内毒素血症相互作用,我们进一步研究了在 RS 和 CS 干预后被来自人类捐赠者的微生物群定植的小鼠的体内肠道通透性。禁食6小时后,给小鼠灌胃异硫氰酸荧光素标记的葡聚糖(DX-4000-FITC),随后通过尾静脉收集血清样本。使用荧光分光光度计测定血清中DX-4000-FITC的浓度。与定植有CS微生物群的小鼠相比,定植有RS微生物群的小鼠的肠道通透性显着较低(图5c)。
肠道通透性受肠道紧密连接的调节,可阻止病原体和细菌产物的入侵7。我们评估了 RS 诱导的肠道微生物群变化对肠道中主要紧密连接蛋白、闭锁带蛋白 (ZO-1) 和 occludin 表达水平的影响。与接受CS微生物群的小鼠相比,接受RS微生物群的小鼠中ZO-1和occludin基因的表达均显着增加(图5d-f)。此外,在RS干预后,在被来自供体的肠道微生物群定植的小鼠中,肠道屏障的恢复导致肠系膜脂肪组织和循环中的LPS水平显着降低(图5g,h)。这些结果表明,改变的微生物群的有益作用之一是通过保护肠道屏障来阻止 LPS 渗透。
RS 改变的肠道微生物群减少脂质吸收
我们的临床研究表明,与食用 CS 相比,食用 RS 后参与者的 ANGPTL4 和粪便脂质循环水平显着增加。为了研究 ANGPTL4 的增加是否受到 RS 微生物群的调节,我们测量了FMT 实验中小鼠回肠中ANGPTL4的表达水平。与定植有CS微生物群的小鼠相比,定植有RS微生物群的小鼠具有显着更高的回肠ANGPTL4表达(图5i)。ANGPTL4 是脂蛋白脂肪酶 (LPL) 和胰腺脂肪酶的内源性抑制剂,其功能是主要的肠道脂肪酶32。
我们发现,与接受CS微生物群的小鼠相比,定植有RS微生物群的小鼠的肠腔脂肪酶活性显着降低(图5j)。根据受抑制的管腔脂肪酶活性,与CS微生物群定植的小鼠相比,RS微生物群定植的小鼠表现出回肠中较低的TG水平和粪便中较高的TG水平(图5k,l)。与定植有CS微生物群的小鼠相比,定植有RS微生物群的小鼠的循环ANGPTL4水平也显着增加(图5m)。这些结果表明,RS 诱导的肠道微生物群变化能够通过调节肠道 ANGPTL4 来减少脂质吸收。
青春双歧杆菌对肥胖的影响
RS引起的青春双歧杆菌的增加与腹部肥胖的减轻密切相关。鉴于此,我们进一步研究了肠道中的青春 双歧杆菌与传统饲养的小鼠宿主肥胖之间的潜在因果关系。小鼠被随机分为三组,饮用含有活青春 双歧杆菌、热灭活青春 双歧杆菌或盐水的无菌水,持续5周(图6a,b)。用活的青春双歧 杆菌补充饮用水显着减轻了西方饮食喂养小鼠的体重增加(图6c)和肥胖(图6d )。
我们进一步研究了青春 双歧杆菌是否调节肠道中ANGPTL4的表达。与给予补充有热灭活的青春 双歧杆菌的饮用水的小鼠相比,给予补充有活的青春双歧杆菌的饮用水的小鼠的回肠ANGPTL4表达和分泌显着更高(图6e-g)。与我们的临床试验一致,青春双歧杆菌与血清 ANGPTL4 水平呈正相关。接受活青春双歧杆菌的小鼠管腔脂肪酶活性显着降低(图6h)。相应地,这些小鼠的回肠TG降低,粪便TG水平升高(图6i,j)。
在用活青春双歧杆菌治疗的小鼠中,循环ANGPTL4水平也显着增加(图6k)。我们的研究结果表明,RS 诱导的关键物种之一青春双歧杆菌可能通过影响肠道 ANGPTL4 来保护小鼠免受饮食诱导的肥胖。此外,RS干预后参与者的FGF21水平显着下降,表明FGF21敏感性提高(图1m)。我们发现,用青少年双歧杆菌治疗小鼠,通过抑制 LPS-TLR4-NF-κB 通路,增加了脂肪组织中对 FGF21 的敏感性。
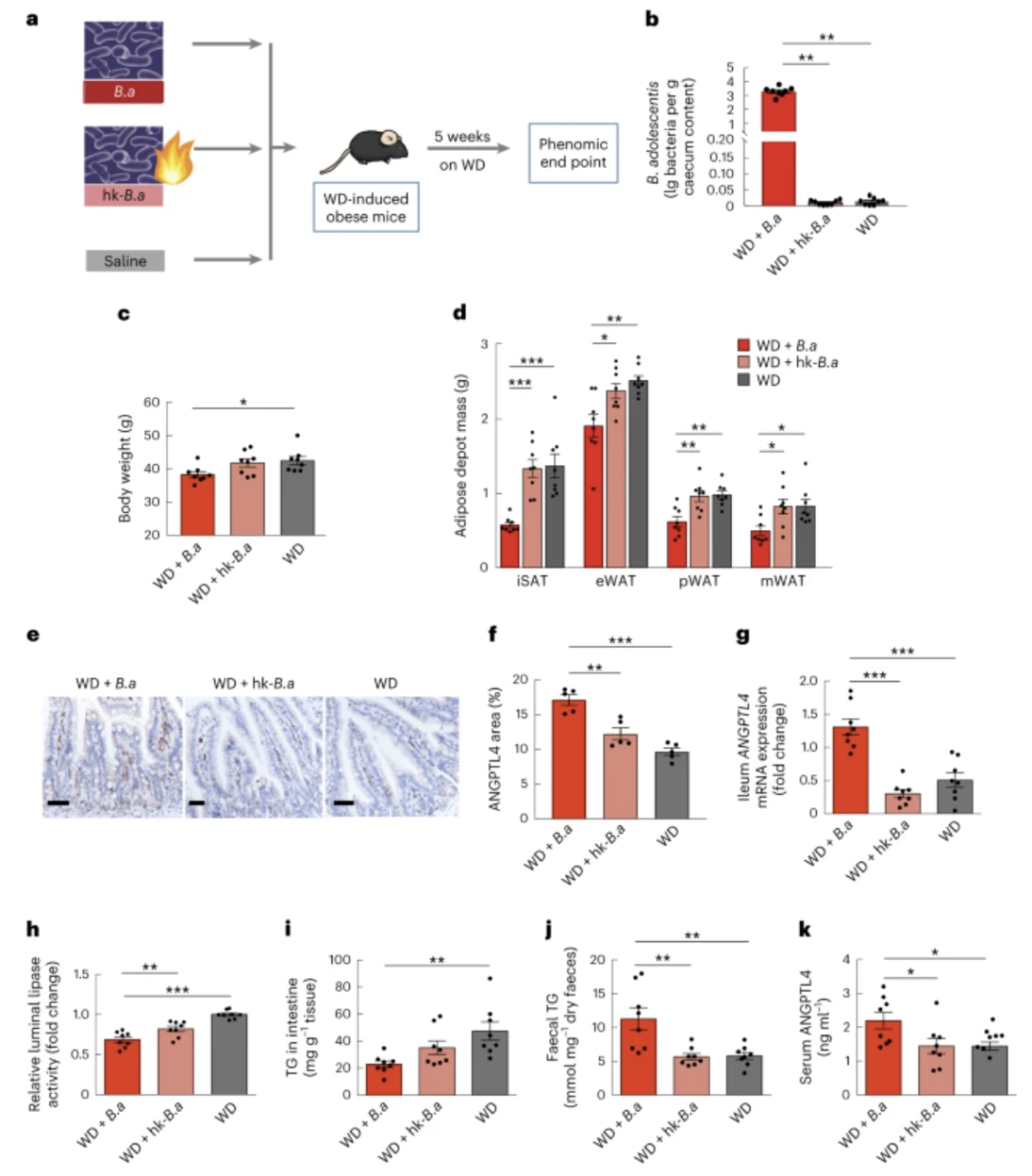
图 6:青春 双歧杆菌可预防饮食引起的肥胖并影响肠道和循环 ANGPTL4。
a ,青春 双歧杆菌(Ba )补充策略示意图。WD诱导的肥胖C57BL/6小鼠补充含有活的(WD+ Ba )或热灭活的青春 双歧杆菌(WD+hk- Ba )或盐水(WD)的饮用水5周。b ,使用三组盲肠内容物中的特异性引物通过 qPCR 定量青春 双歧杆菌的丰度。c – k,治疗5周后三组测量的参数:体重(c)。脂肪库质量包括 iSAT、eWAT、pWAT 和 mWAT ( d )。通过免疫组织化学染色(比例尺,100 µm)观察小鼠回肠中 ANGPTL4 的定位和水平(e)。显示了每组的代表性图像。ANGPTL4 +面积占回肠粘膜总面积的百分比 ( f )。回肠中ANGPTL4 mRNA 表达水平 ( g )。相对肠腔脂肪酶活性(h)。回肠中的 TG 水平 ( i )。粪便 TG 水平 ( j )。血清 ANGPTL4 水平 ( k )。 每组n = 8 个生物学重复( b – d,g – k)。数据在三个独立实验中重现。数据以平均值±sem 表示。显着性通过单向方差分析(正态分布)、Tukey 事后检验或 Kruskal-Wallis 检验(非正态分布)、Dunn 检验来确定。* P = 0.04 ( c )、0.02、0.04、0.03 ( d ) 以及 0.05 和 0.05 ( k )。** P = 0.001、0.004 ( b )、0.003、0.004 和 0.002 ( d )、0.001 ( f )、0.009 ( h )、0.002 ( i ) 以及 0.003 和 0.003 ( j )。*** P < 0.001。
我们进一步对用青春 双歧杆菌治疗的常规饲养小鼠进行了靶向代谢组学分析(扩展数据图9a-c)。青春 双歧杆菌干预后,初级胆汁酸,包括牛磺-β鼠胆酸(TβMCA)、牛磺鹅去氧胆酸(TCDCA)和牛磺胆酸(TCA)显着降低,次级胆汁酸牛磺石胆酸显着升高(扩展数据图9a)。关于 SCFA,在青春 双歧杆菌干预后,粪便中异丁酸和戊酸的水平也显着下降(扩展数据图9b)。此外,在青春双歧杆菌干预后,血清支链氨基酸(BCAA)(包括缬氨酸和亮氨酸)的水平显着下降( 扩展数据图9c)。
RS 部分通过青春 双歧杆菌促进体重减轻
为了探讨RS的影响是否取决于肠道微生物群,我们对无菌小鼠进行了RS和CS干预(图7a)。雄性无菌小鼠喂食 C35 饮食(35% 碳水化合物来自 20% RS 或 20% CS)10 周。同时,这些小鼠接受 PBS 或活青春 双歧杆菌治疗8 周。由于RS喂养后肠内容物和管腔厚度显着增加,因此通常用剖腹体重来评价减肥效果。与传统环境下RS对饮食诱导的肥胖的保护作用不同33,与无菌小鼠中的CS治疗相比,RS治疗对去内脏体重、葡萄糖耐量和胰岛素敏感性没有显着变化(图7c-f)。
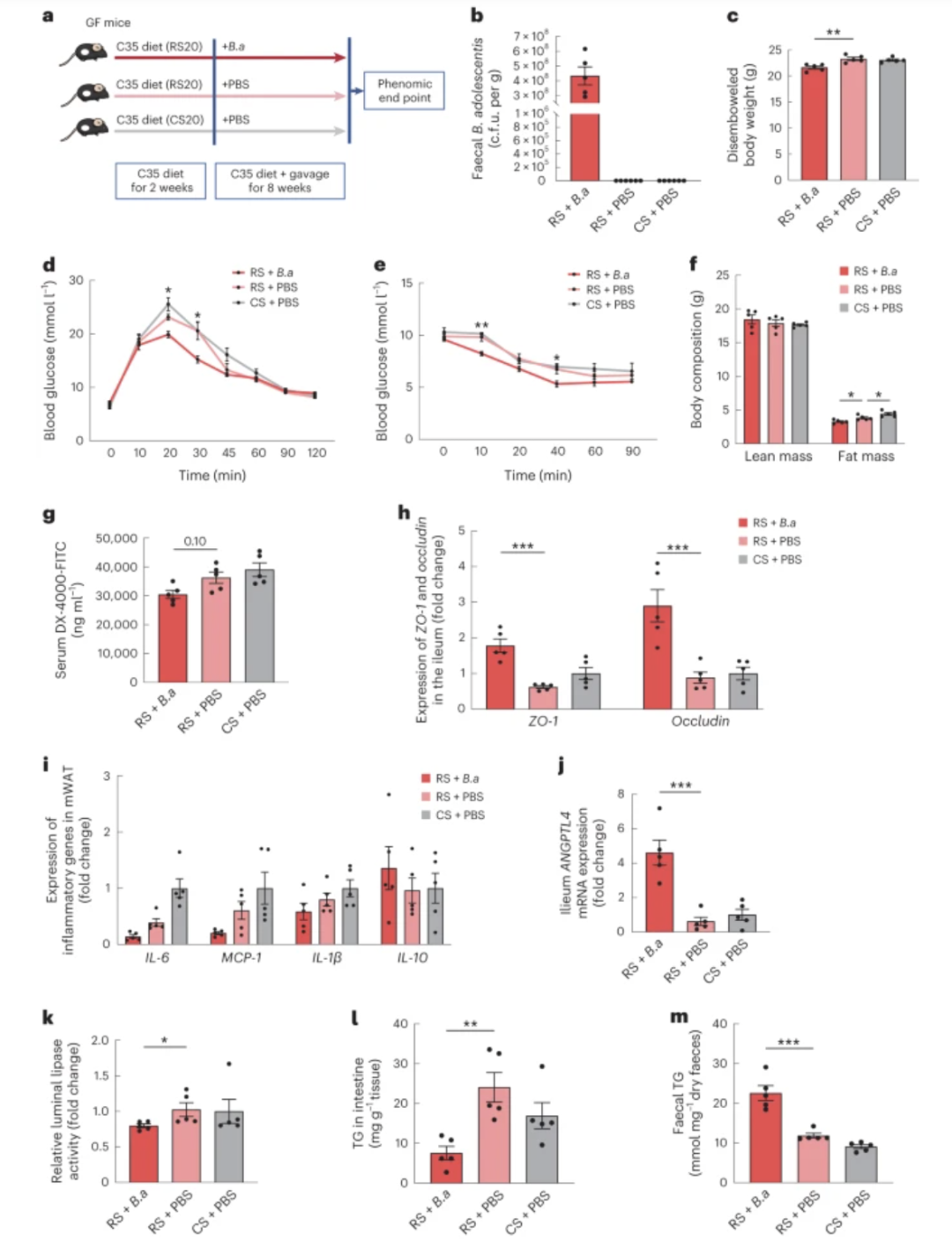
图 7:RS 对有和没有青春双歧 杆菌的无菌小鼠的影响
a ,青春 双歧杆菌(Ba )补充策略示意图。将青少年双歧杆菌 或 PBS 口服接种到无菌 (GF) 小鼠体内,小鼠饮食中含有 20% 蛋白质、45% 脂肪和 35% 碳水化合物(20% CS 或 20% RS,其余 80% 来自麦芽糖糊精),持续 8 周。b,青春双歧杆菌的丰度量化为盲肠内容物中的总平板计数。cfu,菌落形成单位。c,剖腹体重。d,腹膜内葡萄糖耐量试验(GTT)的葡萄糖偏移曲线。e,腹膜内胰岛素耐量试验(ITT)的葡萄糖偏移曲线。f,身体质量成分。g ,补充青春双歧杆菌8周后,通过口服管饲后1小时测量DX-4000-FITC的血清浓度来确定体内肠道通透性。h,回肠中ZO-1和occludin的表达。i,mWAT 中炎症基因的表达。j ,回肠中ANGPTL4的mRNA表达水平。k,相对肠腔脂肪酶活性。l,回肠中的TG水平。m,粪便TG水平。数据为平均值±sem(n = 每组 5 个生物重复)。* P = 0.04 和 0.03 ( d )、0.02 ( e )、0.05、0.04 ( f ) 和 0.03 ( k ), ** P = 0.002 ( c )、0.002 ( e ) 和 0.004 ( l ),*** P < 0.001 基于单向方差分析(正态分布),然后是 Dunnett 检验或 Kruskal-Wallis 检验(非正态分布),然后是 Dunn 检验。*表示RS+ Ba和RS+PBS( d , e )之间的比较 。
为了进一步研究有和没有青春 双歧杆菌的 RS 的影响,我们用 RS 和青春 双歧杆菌一起喂养无菌小鼠。与未添加青春 双歧杆菌的小鼠相比,这些小鼠的去内脏体重和脂肪量显着下降,并且葡萄糖耐量和胰岛素敏感性得到改善(图7c-f)。青春双歧 杆菌降低了无菌小鼠的肠道通透性并增加了肠道ANGPTL4的产生,这与我们在常规环境中的发现一致(图7g-k)。灌胃青春双歧杆菌的无菌小鼠表现 出管腔脂肪酶活性和回肠TG水平降低,同时粪便TG水平升高(图7l,m)。这些结果表明肠道微生物群对于 RS 预防饮食引起的肥胖的作用至关重要。RS 至少部分地通过青少年双歧杆菌 通过恢复肠道屏障和调节肠道中 ANGPTL4 的产生来减少炎症,从而阻碍脂质吸收,从而在促进体重减轻方面发挥了作用。
此外,无菌小鼠血清中初级胆汁酸水平高于常规环境,而次级胆汁酸水平相对较低34。GCA(一种结合胆汁酸)和 DCA(一种次级胆汁酸)在人类进行 RS 干预后表现出增加的趋势。无菌小鼠经青春双歧杆菌干预后 , GCA 和 DCA 显着增加。无菌小鼠经青春 双歧杆菌干预后,初级胆汁酸,包括TβMCA、TCDCA和TCA显着下降(扩展数据图9d)。青春双歧杆菌干预后,血清支链氨基酸和粪便异丁酸水平下降(扩展数据图9e,f)。
03
分析
这项安慰剂对照、双盲和交叉设计的试验是在体重超标的个体中进行的。
作者发现,补充 RS 与等能量和均衡饮食相结合,可以显着减轻人类体重并提高胰岛素敏感性。RS 治疗重塑了微生物组结构并改变了代谢物。肠道微生物群在 RS 的减肥功效中发挥着关键作用。青春 双歧杆菌在小鼠中的单定植与RS对人类的益处有关,可以防止小鼠因饮食引起的肥胖。从机制上讲,RS诱导的肠道微生物群变化影响胆汁酸代谢,通过肠道屏障恢复减少炎症,通过调节ANGPTL4抑制脂质吸收并提高FGF21敏感性。RS 至少部分通过青春 双歧杆菌在促进体重减轻方面发挥了作用。具体来说,这项研究提供的证据表明,RS(40 g d -1,2型)作为膳食补充剂持续8周可以帮助体重超重的个体实现减肥。
在这项喂养研究中,作者在整个研究过程中为参与者提供了背景饮食。正如 CS 治疗后观察到的,等能量且平衡的背景饮食在 8 周内不会影响体重或脂肪含量。
在之前使用 RS 的试验中,相对不足的 RS 摄入量(脂肪:纤维比例 ≈53:27 g d −1)35或高脂肪摄入量(脂肪:RS 比例 ≈77:40 g d −1)导致胰岛素抵抗或脂肪减少没有体重减轻的百分比。其他试验没有规定背景饮食,脂肪摄入量要么太高,要么没有评估。
高脂肪饮食会引起肠道微生物群的变化,从而导致肠道健康受损。在基于微生物组的疗法中,背景饮食对于疗效至关重要。在啮齿类动物中,补充 RS 的低脂饮食(18% 能量)对宿主具有有益作用,但高脂肪饮食(42% 能量)会减弱 RS 发酵和有益作用。
由于40 g d -1 RS被认为是相对较高的剂量,可以在没有副作用的情况下输送,因此日常饮食中的脂肪含量对于RS的效果很重要。我们的研究提供了使用 RS 作为补充剂的有效饮食建议(40 g d −1含有 25-30% 脂肪的均衡背景饮食),这可能有助于实现显着的减肥效果。背景饮食的设计和良好的依从性使我们能够减轻已知对肠道微生物组和代谢组有影响的显着混杂变量的影响。
这项研究是报告 RS2 干预后特定微生物群特征的少数研究之一。RS2消耗后通常富集的分类群包括R. bromii、B.teenis 、Faecalibacter prausnitzii和Eubacterectale。作为主要 RS2 降解剂的R. bromii在之前的 RS2 干预措施中得到富集。在这项研究中,对体重超标的个体进行 RS 干预后,R. bromii和B.青春期显着增加。青少年双歧杆菌丰度的增加与 BMI 和 VFA 的降低密切相关,表明其在 RS 的减肥功效中发挥着重要作用。
基线时肠道微生物组中含有青春双歧杆菌的参与者在 RS 治疗后表现出更大的脂肪量下降。此外,我们通过人类微生物群植入传播了RS对小鼠宿主肥胖和葡萄糖代谢的有益影响,这强化了我们的假设,即RS诱导的微生物群变化可以驱动有益的宿主结果。此外,肠道微生物群对于 RS 的益处至关重要,这一点通过对无菌小鼠进行 RS 干预得到了证实。
微生物群衍生代谢物的产生参与益生元的功能。胆汁酸是将肠道微生物群与宿主联系起来的重要信号代谢物。其中,次级胆汁酸GDCA在RS处理后显着增加,其他次级胆汁酸,例如DCA、7-ketoLCA和TDCA,表现出增加的趋势。据报道,次级胆汁酸可以改善肝脏脂肪变性并增强胰岛素敏感性。BSH 进行胆汁酸解离。在我们的研究中, BSH基因的丰度下降,并与B. vulgatus显着相关。抑制 BSH 活性成为调节脂质和能量稳态的潜在有益方法,导致结合胆汁酸增加。在我们的队列中,结合胆汁酸,包括 GCA、GDCA 和 TDCA,在 RS 治疗后表现出增加的趋势。
与人类的研究结果一致,青春双歧杆菌对无菌小鼠进行干预后,GCA 和 DCA 显着 增加。GCA 和 DCA 分别是 FXR 和武田 G 蛋白偶联受体 (TGR5) 的激动剂,它们是调节葡萄糖、脂质和能量代谢的已知受体。在常规环境和无菌环境中,青春双歧杆菌干预后, 血清支链氨基酸(包括缬氨酸和亮氨酸)水平均显着降低。
对患有非酒精性脂肪肝病 (NAFLD) 的参与者进行 RS 干预后,支链氨基酸的血清水平下降。粪便异丁酸是缬氨酸蛋白水解的微生物产物,在我们的研究中进行 RS 干预后减少。
能量调节和炎症之间的相互作用受到肠道微生物组的调节,影响肠道通透性并产生促炎因子,对代谢健康表现出不同的影响。肠粘膜的完整性通过细胞间紧密连接来维持,细胞间紧密连接是肠道通透性的重要调节剂。在接受 RS 诱导的微生物群的小鼠中,两种紧密连接蛋白 occludin 和 ZO-1 的水平在回肠中升高,而循环 LPS 降低,这表明 RS 改变的肠道微生物群是保护肠道屏障的另一种机制。
慢性低度炎症和免疫系统激活与肥胖和胰岛素抵抗的发展有关。受肠道微生物群影响的体重管理也可能通过调节参与能量利用和稳态的分泌蛋白来实现。值得注意的是,宏基因组学分析表明,RS 治疗后青春双歧 杆菌丰度的增加与血清 ANGPTL4 呈正相关。ANGPTL4 与肠道微生物群的关系引起了人们的极大兴趣。与传统饲养的小鼠相比,无菌小鼠的肠道ANGPTL4表达升高。ANGPTL4 水平的降低通过增加脂肪组织中的 LPL 活性,有助于增加常规化时的脂肪积累。
在无菌小鼠中定植产乙酸盐的多形拟杆菌或产甲烷的史密斯甲烷短杆菌,导致肠道ANGPTL4表达降低,而产丁酸盐的细菌酪丁酸梭菌定植显着升高肠道ANGPTL4表达。对此结果的一个可能的解释是,肠道 ANGPTL4的调节取决于肠道微生物群的组成,肠道微生物群的组成决定了所形成的微生物代谢物的混合物以及底物供应。使用 FMT,我们观察到 RS 诱导的肠道微生物群变化通过调节肠道 ANGPTL4 减少脂质吸收。此外,在常规和无菌环境中饲养的饮食诱导的肥胖小鼠中,青春双歧杆菌的定植显着升高了肠道ANGPTL4水平。
因此,通过 ANGPTL4(脂质吸收的关键调节因子)通过肠道微生物群调节来控制胰腺脂肪酶活性,可能是改变体内脂肪储存的关键方法。青春期双歧杆菌 补充剂可以通过增强肝脏中 FGF21 的敏感性来减轻 NAFLD。我们的结果还表明,青春双歧 杆菌治疗通过抑制 LPS-TLR4-NF-κB 通路来增加脂肪组织中 FGF21 的敏感性。FGF21 类似物和模拟物的多项临床试验显示,血脂状况得到改善,脂联素增加,体重减轻。研究发现,逆转肥胖诱导的脂肪组织中 FGF21 抗性可以作为治疗肥胖及相关疾病的替代方法。
该研究的主要局限性是样本量相对较小,并且参与者的纳入标准严格,限制了结果的普遍性。我们的依赖于数据库、基于分类单元的分析虽然为分类分配提供了全面且注释良好的信息,但也面临着局限性,例如丢弃难以分类的序列和忽视菌株水平的功能多样性。我们承认,整合宏基因组组装的基因组可以增强我们对微生物生态学的理解,并有可能改善用于临床转化的稳健生物标志物的识别。我们的结果具有更广泛的普遍性,需要在更大、更多样化的群体中进行额外的验证。未来的研究需要特别关注 RS 补充剂中微生物群的个体动态和功能反应。
进一步研究有必要深入了解 RS 改变的胆汁酸和肠道微生物群之间的串扰及其对宿主代谢的影响。尽管对其他人群的适用性尚未得到证实,但我们的研究提供了证据,表明 RS(40 g d -1,2 型)作为膳食补充剂有助于体重超标的个体减肥。我们的研究还提供了有效的饮食设计和 RS,以减轻影响肠道微生物组的主要混杂变量的影响。RS 驱动的肠道微生物群变化对其效果至关重要。肠道微生物群在这种减肥机制中发挥着关键作用,可能通过与低度炎症的相互作用以及能量平衡相关分泌蛋白的调节来调节肥胖。通过交叉设计,我们发现体重在洗脱期得到了恢复。
体重反弹是减肥治疗的最大挑战之一,这种情况发生在停止药物治疗(例如 GLP-1 激动剂)后,甚至在减肥手术后49。长期坚持富含RS的饮食模式以维持微生物组的组成可能对于维持体重至关重要。由于 RS 天然存在于食物中,也可以添加到日常饮食中,因此我们的研究结果为治疗肥胖及其相关代谢紊乱提供了一种务实的生活方式。
通过饮食控制肠道微生物组成可能是改变宿主能量平衡以促进健康的一种策略。


